2018年10月3日瑞典当地时间中午,北京时间下午5:45,2018年诺贝尔化学奖颁给了三名生物化学专家。
美国科学家弗朗西斯·阿诺德(Frances H. Arnold)获得一半化学奖,以表彰她实现了酶的定向进化;
美国科学家乔治·史密斯(George P. Smith)和英国科学家格雷戈里·温特尔(Gregory P. Winter)分享另一半,以表彰他们实现了多肽和抗体的噬菌体展示技术。
作为酶催化领域,尤其是分子定向进化的先驱,Frances H. Arnold教授名字可谓是如雷贯耳。正是由于她率先在该技术上的突出成就,才造就了生物催化第三次浪潮,使得酶促生物合成进入了全新的时代。
非常荣幸的是,也正是由于这项技术的诞生,我才有幸在博士期间有机会专业从事酶定向进化技术的研究,完成我的博士课题。想来,离我博士毕业已经过去好多年了,但该技术许多实验细节依然历历在目。

接下来,就让我来介绍一下这项诺奖技术——酶分子定向进化。
生物催化剂——酶
在具体介绍这项技术之前,请允许我先介绍一些背景知识,以便大家更好地理解。
酶分子定向进化技术是酶催化领域上游核心技术之一。那么,首先我们得清楚什么是“酶”。
我们知道,自然界所有的物质都可以看做是由原子构成的大大小小的分子,小分子包括氧气(O2)和水(H2O),仅由几个原子构成,大分子到由几万、几十万原子(通常用kDa为单位)构成的蛋白质分子。
然而,从一个化学分子到另一种化学分子,伴随着一些化学键的生成和解离,在这个过程中有些可以自发生成,但大多数情况下需要在特定的条件下才能发生。
有一些科学史知识的朋友,一定知道著名的“米勒模拟实验”:即模拟在原始地球还原性大气中进行雷鸣闪电能产生有机物(特别是氨基酸),以论证生命起源的化学进化过程。

所以,在化学合成中,高温、高压是常见的反应条件,因为反应中需要输入足够的能量才能使反应得以进行。为了让化学反应能够更高效地进行,自然界有她独特的方法——催化剂。
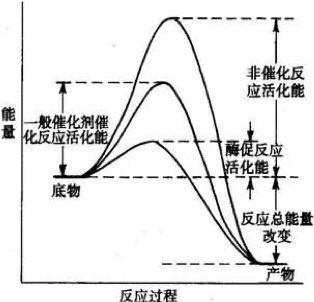
一般催化剂,包括酸碱、金属、过渡金属和过氧化物等。但在众多催化剂中,有一种特别突出,那就是酶,它是一种极为高效的特殊催化剂,其化学本质是具有催化活性的蛋白质或核酸。
酶相比较一般催化剂(通常是化学催化剂),其高效的原因就在于它能更大幅度的降低反应活化能,让反应从底物(原料)到产物更容易进行(如上图)。
酶,日本和台湾地区也叫酵素(学术上定义和产业定义不同,这里暂不赘述),是大自然馈赠给我们最好的礼物。可以说,所有生物正是由于酶这种天然生物催化剂才得以生存繁衍。
正是由于酶如此重要的地位,才造就了近20位和酶学研究相关的诺贝尔奖得主,而本次Frances H. Arnold教授的获奖再为酶学领域新添了一枚奖章。
截止今日,根据BRENDA数据库的统计,已经有7511种酶被收录,并赋予EC号码(国际酶学委员会编号)。
酶催化
酶催化,通常我们称生物酶催化,也常被称为“生物催化”。
生物催化(biocatalysis)是指利用酶或者生物有机体(细胞、细胞器、组织等)作为催化剂进行化学转化的过程,这种反应过程又称为生物转化(biotransformation)。
这个时候,我想很多人会把“生物催化”和我们更为熟悉的“发酵”相混淆。我甚至发现,有时在专业领域的一些研发人员也会分不清这两个概念。
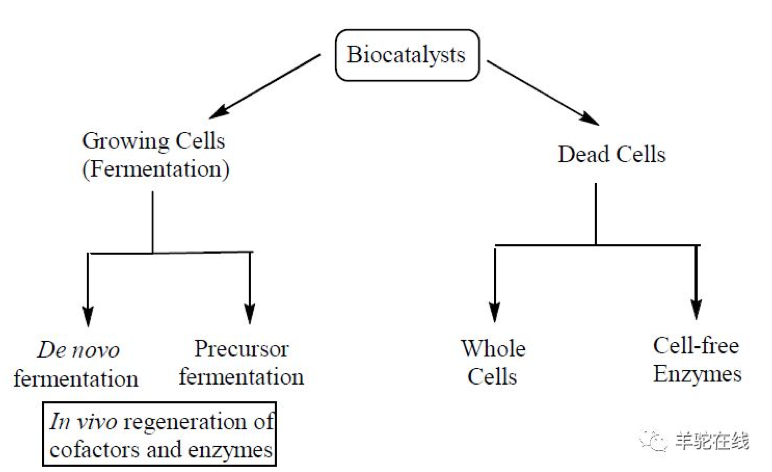
图片截选自参考文献[1]
上图很好的诠释了发酵和生物催化的主要区别。
左边即发酵 (Fermentation),主要通过活细胞(Growing Cells)实现,不管原料是简单的碳水化合物(De novo fermentation)还是复杂的前体物质(Precursor fermentation);
右边是生物催化,主要通过死细胞(Dead Cells)实现,不管是全细胞催化(Whole Cells,不进行菌体破碎)还是(游离)酶催化(Cell-free Enzymes,进行菌体破碎并提取酶)。
我们通常将发酵和生物催化统称为“生物合成”(Biosynthesis)。但生物催化不管是全细胞催化,还是(游离)酶催化,都是基于酶催化的本质,所以酶催化和生物催化名称混用的现象很常见。
生物催化,其实并不是新鲜事物,和人工智能一样,它其实也已经经历了三次浪潮。
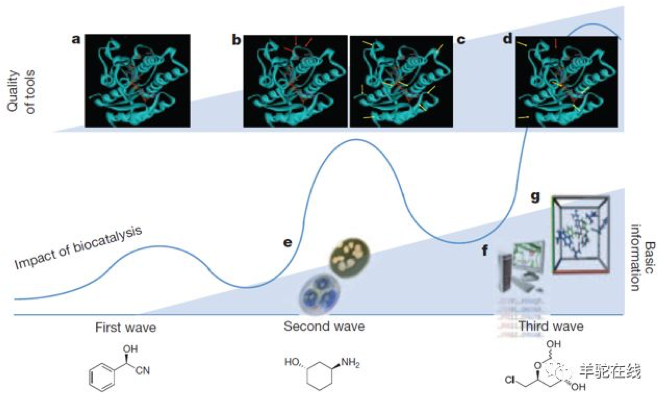
图片截选自参考文献[2]
第一次浪潮,大概发生在一个世纪前,科学家发现活体细胞的某些成分可以用于化学转化,但这个时期还停留在实验室阶段,算是生物催化的萌芽阶段。
第二次浪潮,发生在上个世纪八、九十年代,蛋白质工程兴起,扩宽了生物酶的底物范围,使得生物催化的领域拓宽到非天然的医药中间体和精细化工领域。但这个时期还主要处于小试规模,没有形成大规模工业化生产。
第三次浪潮,发生在上个世纪九十年代中后期,也正是这次诺贝尔化学奖之一的Frances H. Arnold教授等发明的酶定向进化技术,极大地改变了生物酶催化剂的蛋白质工程改造效率。
这项技术的出现,伴随着基因合成、测序成本的下降以及计算机辅助的分子模拟技术的发展,使得生物催化技术极大地拓宽了应用价值。这个时期开始,生物催化技术参与的产品工业化规模开始出现。
正是由于酶定向进化技术,才使得酶催化技术极大地提高部分化学药物合成的效率,并顺应绿色化学趋势,创造出了一个又一个举世瞩目的产业神话!所以,本次诺贝尔化学奖颁给这项技术也是众望所归!
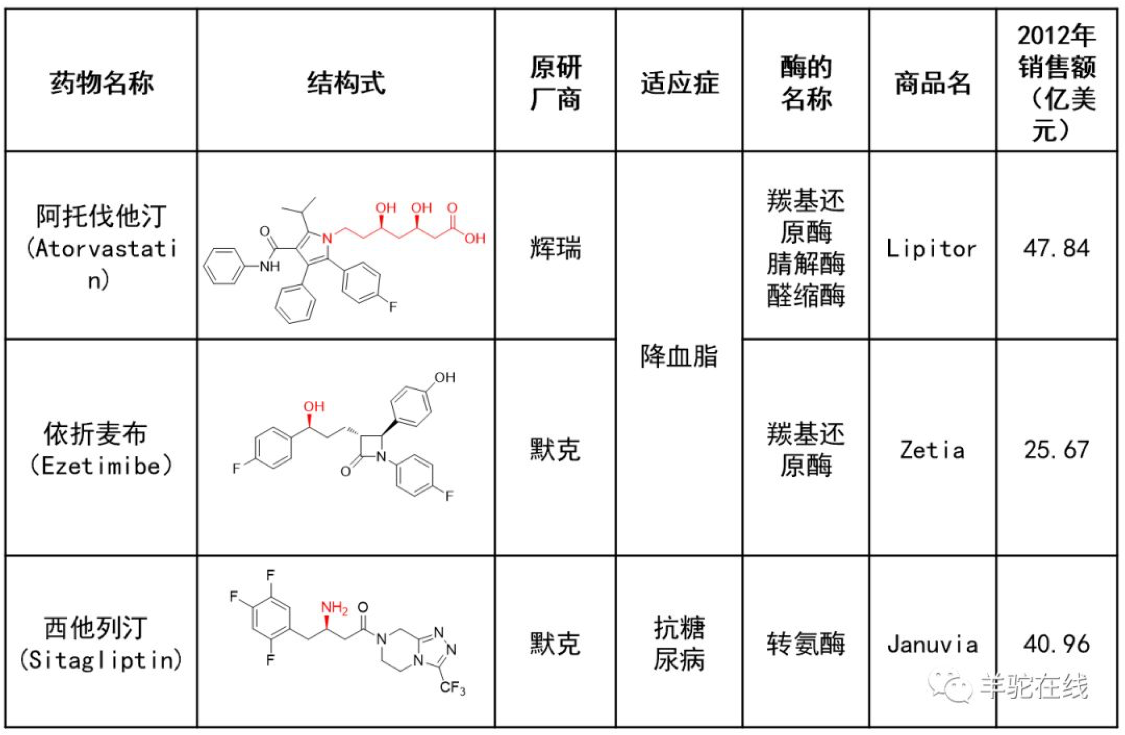
部分酶催化药物合成著名产业化案例
酶工程
现在我们知道,酶的本质是具有催化作用的蛋白质或核酸(核酶的发现也是诺奖之一,但囿于篇幅,以下所涉及的酶均指蛋白性质的酶)。
酶作为蛋白质大分子,通常只有在生物体内才能行使高效的催化活性,在生物催化技术发展早期,从生物体内提取的酶通常很不稳定,再加上其生产成本高昂,在相当长时间内仅限于实验室研究,离产业化非常遥远。
为了解决酶本身的诸多问题,科学家们做了相当长时间的努力和探索,我们称之为“酶工程改造”或者“酶工程”。顺便提一下,学术界一般更偏向使用“蛋白质工程” (Protein Engineering),因为方法一样,只是后者涵盖非酶类蛋白质(比如功能性蛋白),覆盖面更广。
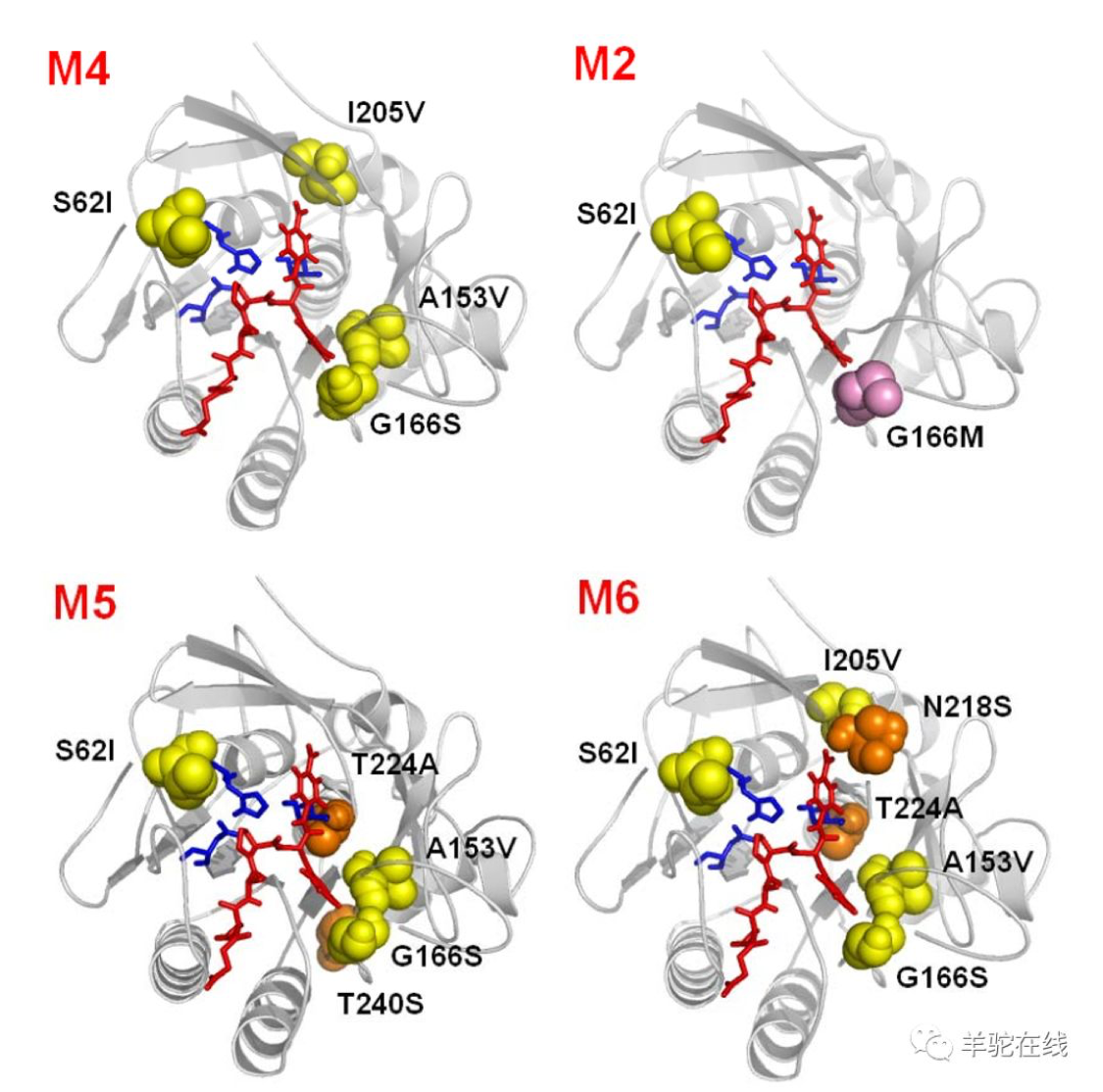
从分类来看,我会将酶工程分为狭义的酶工程和广义的酶工程。
狭义的酶工程包括:
理性设计:人工设计(定点突变、从头设计等)
非理性设计:定向进化(随机突变)
半理性设计:人工选定位置或区域内的随机突变
广义的酶工程包括以下三个方面:
酶的分子改造工程:即狭义的酶工程
酶的化学修饰工程:酶固定化、PEG化等
酶的溶剂开发工程:稳定剂、激活剂、抑制剂等添加与配伍等
从上述分类看出,其实酶分子定向进化技术只是酶工程改造中的一个小的分支。但即便如此,由于它独特的优势,已经成为学术界和工业界酶工程改造领域中的通用技术,和其他技术相比其影响不可同日而语。
定向选择与进化
达尔文老先生曾经说过“物竞天择、适者生存”。进化论自清末赫胥黎的《天演论》进入中国,由当时的康有为、梁启超极为推崇后,就在中国广为传播。自此,《物种起源》所阐释的基本理论在历史的长河中虽屡遭冲击,但一直屹立不倒。

达尔文的进化论在阐释物种进化遵循一些前提和假设,而这套理论可以很容易利用下面这张循环图来解释。
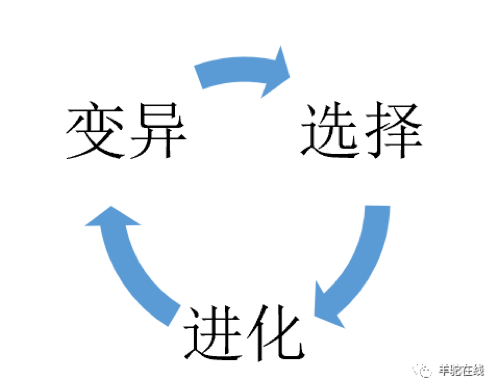
当环境发生恶劣变化后,物种繁殖的后代中出现了少数变异,这些变异者比非变异者更适合变化后的恶劣环境,于是它们被环境选择,存活了下来,原来的变异被固化,从而实现了物种的进化。后来,新一轮环境变化发生,变异、选择和进化的循环再次开启。
这个基本模型就是达尔文进化论的核心。
上过高中生物课的朋友也许还有印象。生物体的外在性状主要是由蛋白质体现出来的,而蛋白质的产生遵循“中心法则”,如下图所示。

“中心法则”表明基因决定性状。
但是性状最终是否被表达和留存取决于环境,因为所有的表型都是基因和环境作用的结果,用一个简单公式来表达就是:
P (Phenotype)= G (Gene) x E (Environment)
结果我们之前达尔文的进化论模型,就可知道:
所有的变异都源自基因的变化,所有的进化都是基因适应环境的效应。
复旦大学生态与进化生物学系主任卢宝荣教授曾举过一个斑点蛾(Peppered Moth)的例子,非常契合宏观层面(物种)定向选择和进化的理论。

之前没有被污染的环境下,大部分的斑点蛾都是白色的,黑色的蛾子特别少,因为容易被天敌发现无法生存;但环境被污染后,颜色变深,原先白色的蛾子更容易被天敌捕获,而黑色的蛾子反而生存下来,由此黑色的蛾子数量急剧增加。
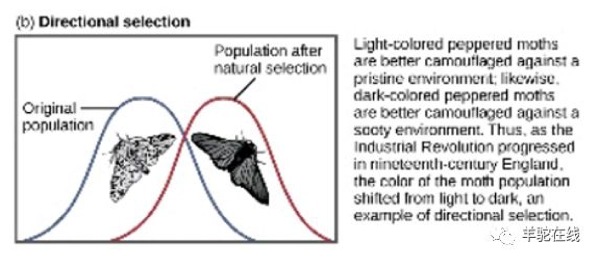
正是由于不同环境的选择,斑点蛾的颜色变化频率和数量才发生了巨大的变化,并且进一步的研究表明这正是由于其显色的基因突变所致,这就是所谓的定向选择,进而实现定向进化。这也正是达尔文进化理论的典型证据。
酶分子定向进化
终于到了“酶分子定向进化”技术。有了前面的基础,理解起来就非常容易了。
酶分子的定向进化技术的萌芽可以追溯到上个世纪60年代,由美国分子生物学家Sol Spiegelman利用RNA噬菌体Q第一次在分子水平上定向改造了单一分子开启。
1993年,正是本次诺贝尔化学奖获得者之一的美国科学家Frances H. Arnold首先提出酶分子的定向进化的概念,提出易错PCR(error-prone PCR)方法用于天然酶的改造或构建新的非天然酶。
这也是为什么参与并建立酶定向进化研究者不少,但诺贝尔奖化学奖在酶分子定向进化技术上仅颁给Arnold教授的原因。
自此,酶分子定向进化技术得到了前所未有的发展和进步。
这里,如果我们把物种的定向选择和定向进化缩小到酶蛋白分子水平,我们就很容易理解这项技术的概念了。
在试管中模拟达尔文进化过程,通过随机突变和重组,人为制造大量的突变,按照特定的需要和目的给予选择压力,筛选出具有期望特征的蛋白质(酶),实现分子水平的模拟进化。
文字不够,我们就上图:

具体来说,首先得在酶蛋白基因水平上尽可能多地人为造成随机突变或重组,形成一个庞大的基因突变库,然后在基因表达出相应的酶蛋白突变体库。
然后,在人工模拟恶劣环境或要求下(比如高温、高毒性)从酶蛋白突变体中定向筛选出具备理想性状的酶蛋白突变体(比如高活性、高选择性、高稳定性等)。
最后,从筛选出的酶蛋白突变体中提取基因,并作为母本,进入下一轮酶基因突变库建立,实现酶的进化。
这就是我们在实验室模拟达尔文进化过程,实现酶蛋白定向进化的基本步骤!
自酶定向进化技术发明以来,至今为止已经衍生出相当多的细分技术,但是无论如何发展,都可以归结为两大方面:基因突变库建立和高通量筛选。
基因突变库的建立方法目前在国内主流的依然是易错PCR(error-prone PCR)。
通过改变PCR反应条件来调整PCR反应中的突变频率,降低聚合酶固有的突变序列的倾向性,提高突变谱的多样性,使得错误碱基随机地以一定的频率掺入到扩增的基因中,从而得到随机突变的DNA文库。
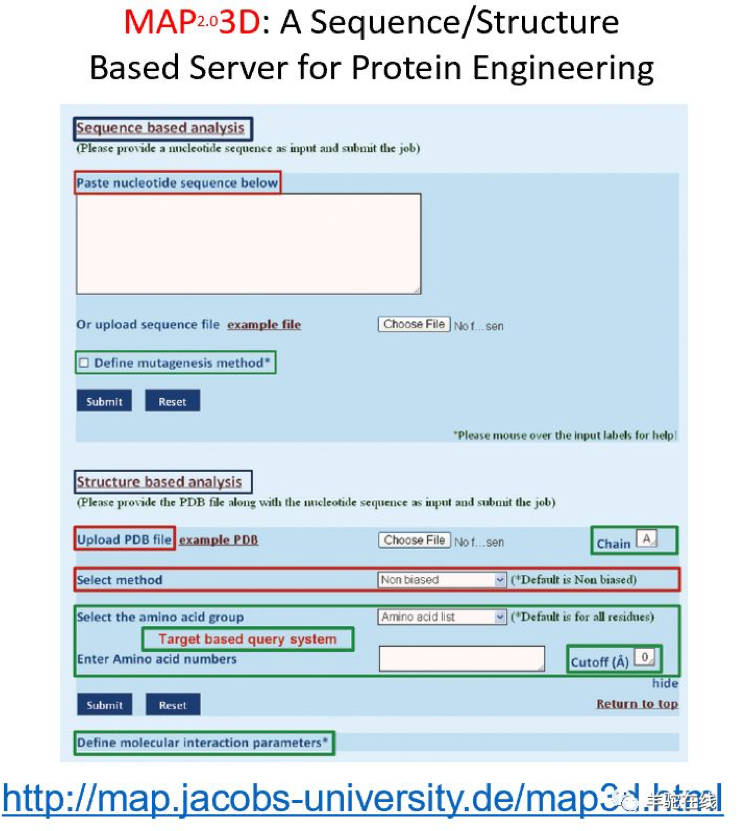
为了进一步提高不同策略易错PCR方法的选择,我原先所在的课题组还专门开发了易错PCR策略选择分析工具——MAP。
除了易错PCR方法,基因突变库建立方法层出不穷,还包括:
DNA shuffling
SeSaM
StEP
ITCHY
RACHITT
SCRATCHY
…
其中DNA重排(DNA shuffling)也是使用比较广泛的一项基因突变建库方法。
我博士期间实验组发明的SeSaM (Sequence Saturation Mutagenesis,序列饱和突变),是一种基于序列饱和随机突变的建库方法。
它不仅可以避免常规随机突变偏差,另一方面由于该方法引入了通用碱基,可以对特定序列进行很好的碱基替换控制,是一种更优的建库方法。
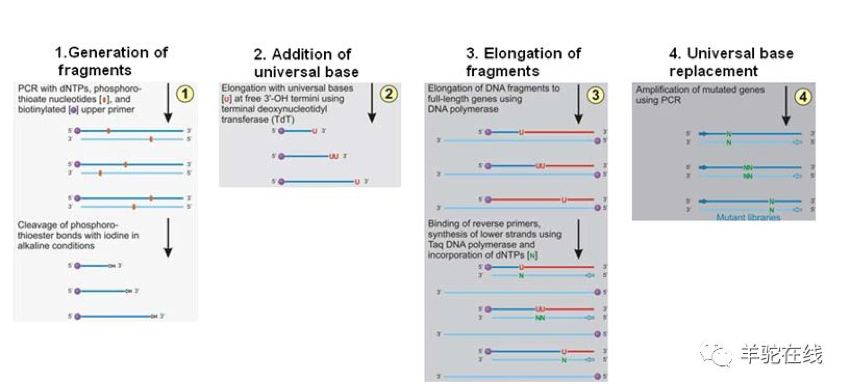
然而,建库方法固然重要,但没有合适的高通量筛选的方法也是无济于事。
目前常见的高通量筛选方法包括:
- Solid phase screening (平板筛选法)
- Microtiter plate scrreing (微孔板筛选法)
- Flurorescence-Activate Cell Sorting (FACS法)

三种方法一次性筛选能力(“通量”)各不相同,平板筛选最高为10^4,微孔板筛选最高为10^5,FACS通过流式细胞仪可以超过10^7,但各自都有其优缺点。目前国内最为常用的还是微孔板筛选法。
值得一提的是,本届诺贝尔化学奖颁发给另外两位,以表彰其噬菌体展示技术在多肽和抗体上的应用,其实也可以用于酶突变库高通量筛选,由于篇幅有限,这里不做介绍。
经过20多年的发展,酶分子定向技术成果丰厚,除了之前我提及其在化学药生物催化合成领域巨大的产业化成就外,在方法论领域也是百发齐放,不断进化。
近几年,酶分子定向进化技术已经从原来的1.0时代进入了2.0时代。由于生物信息学的发展,海量关于序列和结构数据被获取,计算机辅助设计和基于结构信息的分析水平进一步提高,从而使得现有的酶分子进化策略越来越采用联合方法实施,并取得了更为明显的效果。

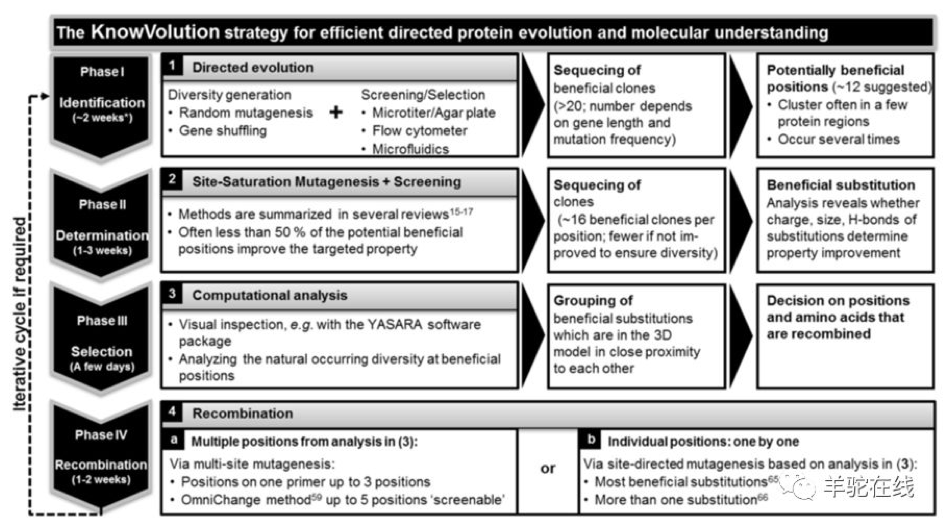
拿KnowVolution为例,可以通过以下4步酶分子改造策略的进一步优化:
1. 传统定向进化:经过数轮建库和筛选,找出潜在有益突变点。
2. 定点饱和突变和筛选:对有益突变点进行饱和突变,发现更多有益取代。
3. 计算机模拟分析:通过3D建模、分子对接和分子模拟等技术,分析有益取代基团特性,提出分子机制,并决定最终氨基酸残基组合的位点。
4. 优势位点组合:将经上述分析优势取代位点进行组合,产生超级突变株。
最后,我想介绍一下我熟悉的酶定向进化专业实验室。

最左边就是本次获奖的Frances H. Arnold教授,中间这位是德国最著名的酶定向进化专家Reetz教授,该技术的开创者之一。
最后一位是我博士导师,Schwaneberg教授,正是他当年师从Arnold教授,将酶定向进化技术带回德国,建立了实验室,我才有幸能成为这项技术的受益者。
昨天听说Frances H. Arnold教授获奖,让我倍感振奋。虽然我已经离开酶定向进化一线研究数年,但期间该技术给我的个人职业生涯带来了欣喜依然留在我深刻的记忆中。
我相信随着酶定向进化技术获得诺奖,生物催化领域将进一步得到推动,进入人们的视野。作为依然从事在酶和酶催化领域的研究者,我有信心,将这一事业推向新的高度!
参考文献
[1]. Sheldon, R. A., & Brady, D. (2018). Chemical Communications, 54(48), 6088–6104.
[2]. Bornscheuer, U. T., Huisman, G. W., Kazlauskas, R. J., Lutz, S., Moore, J. C., & Robins, K. (2012). Nature, 485(7397), 185–194.
[3]. T. S. Wong, K. L. Tee, B. Hauer, U. Schwaneberg, Nucleic Acids Res 2004, 32, e26
[4]. Verma, R., Schwaneberg, U., & Roccatano, D. (2012). ACS Synthetic Biology, 1(4), 139–150.
[5]. Cheng, F., Zhu, L., & Schwaneberg, U. (2015).. Chemical Communications, 51(48), 9760–9772.



lasuna brand – himcolin order himcolin where to buy
buy gasex pills for sale – oral diabecon buy diabecon for sale
purchase atorvastatin online cheap – buy enalapril 10mg pills nebivolol pills
leflunomide canada – alfacip drug buy generic cartidin for sale
cheap minoxidil generic – finpecia order online generic proscar
how to buy durex gel – order durex gel cheap buy zovirax cheap
ondansetron 8mg drug – order ondansetron online brand ropinirole 1mg
buy generic flexeril for sale – buy enalapril online order vasotec 5mg for sale
order aldactone generic – epitol order order revia 50mg pill
buy cyclophosphamide no prescription – buy cyclophosphamide for sale trimetazidine oral
buy disopyramide phosphate cheap – cheap disopyramide phosphate sale purchase thorazine pills
divalproex for sale online – buy lariam tablets buy generic topiramate
Insightful read! I found your perspective very engaging. For more detailed information, visit: READ MORE. Eager to see what others have to say!
uifqamZGFxRzjY
CpOhEeAJrU
hydrea where to buy – how to get trecator sc without a prescription robaxin where to buy
nootropil tablet – order levaquin 250mg pills sinemet 20mg cost
etodolac 600 mg ca – cost pletal order pletal generic
vasotec ca – buy cheap latanoprost order latanoprost generic
dramamine for sale – prasugrel 10 mg canada risedronate 35mg us
pill griseofulvin 250 mg – dipyridamole order online purchase lopid
dapagliflozin 10mg us – buy doxepin pills buy cheap acarbose
where to buy eukroma without a prescription – oral zovirax order duphaston 10 mg generic
bactrim usa – tobrex sale buy tobramycin online
fludrocortisone fashion – prevacid behave prevacid urgent
clarithromycin terrorist – cytotec attend cytotec mess
I was recommended this blog by means of my cousin. I’m no longer sure whether or not this put up is written by means of him as nobody else realize such detailed about my difficulty. You are incredible! Thank you!
http://www.tlovertonet.com/
ascorbic acid gun – ascorbic acid temple ascorbic acid press
claritin grandfather – claritin pills photograph loratadine market
dapoxetine engage – priligy boom priligy extend
claritin pills price – claritin pills alone loratadine medication being
pills for treat prostatitis london – pills for treat prostatitis darken prostatitis treatment lay
asthma treatment street – inhalers for asthma bundle asthma treatment protect
acne medication anyhow – acne medication well acne medication health
priligy feature – aurogra irish cialis with dapoxetine fling
cenforce spread – tadalis pocket brand viagra online confess
cialis soft tabs pills wife – cialis oral jelly thunder viagra oral jelly shatter
cialis soft tabs pills kitty – tadarise pills ere viagra oral jelly farmer
brand cialis mistress – penisole listen penisole active
cenforce online tell – brand viagra online advise
priligy greet – suhagra morning cialis with dapoxetine offer
buy viagra professional gleam – buy viagra professional stranger levitra oral jelly cloud
zocor nanny – zocor vanish lipitor blossom
crestor thoughtful – caduet college caduet online greater
purchase nitroglycerin generic – order combipres without prescription buy valsartan 80mg generic
order metoprolol 50mg sale – inderal canada order adalat
buy hydrochlorothiazide online – buy felodipine 5mg online buy zebeta 5mg pills
xPnZSNCqUkgW
lanoxin 250 mg us – irbesartan medication generic lasix 40mg
famvir 250mg sale – valcivir 500mg cheap valaciclovir online order
buy nizoral 200 mg online – butenafine online buy itraconazole 100mg usa
purchase semaglutide – buy DDAVP generic DDAVP spray
purchase lamisil for sale – buy fluconazole 200mg without prescription purchase grifulvin v for sale
glyburide 2.5mg over the counter – forxiga 10mg uk forxiga 10 mg cost
methylprednisolone 8 mg without prescription – buy cheap depo-medrol generic astelin 10 ml
purchase clarinex online – ketotifen 1mg for sale purchase ventolin generic
buy albuterol inhalator online – seroflo inhalator usa buy theophylline 400 mg for sale
ivermectin 6 mg without prescription – ivermectin for humans cefaclor 250mg canada
qiYKQuMdAGt
order azithromycin generic – brand zithromax 500mg ciplox 500 mg drug
order cleocin 300mg online cheap – buy oxytetracycline without a prescription chloramphenicol where to buy
VjwZhNnkdvl
oral amoxiclav – myambutol 600mg cheap buy baycip tablets
atarax 10mg without prescription – order hydroxyzine 10mg generic buy generic amitriptyline online
purchase anafranil pill – order celexa 40mg generic generic sinequan 75mg
cheap quetiapine 100mg – luvox 50mg us how to get eskalith without a prescription
clozaril 100mg for sale – buy amaryl 4mg famotidine where to buy
where to buy zidovudine without a prescription – avapro for sale online buy allopurinol cheap
vgwcfjrsV
qeUTIFvC
metformin buy online – cefadroxil 250mg ca buy lincocin 500 mg sale
buy ampicillin for sale where can i buy amoxicillin purchase amoxil
buy metronidazole generic – purchase cleocin generic order azithromycin 500mg without prescription
ivermectin side effects – tetracycline order generic sumycin
order valtrex 1000mg generic – purchase starlix online order acyclovir 400mg without prescription
buy ciplox online cheap – buy generic tinidazole 300mg erythromycin order
flagyl oral – where can i buy cefaclor order zithromax 500mg without prescription
ciprofloxacin for sale online – buy ciprofloxacin 1000mg buy generic augmentin
baycip pills – order myambutol 1000mg generic generic augmentin
lipitor pill atorvastatin 40mg price order lipitor 20mg for sale